(Contributo di Daniel Nesta, con i nostri ringraziamenti)
ORMONI VEGETALI
Le piante cormofite possiedono una struttura altamente organizzata. Si definisce cormo il corpo delle piante superiori, organizzato nei tre organi fondamentali: radice,fusto o caule, foglia. Nel cormo le cellule presentano spiccate differenziazioni morfologiche e funzionali nell'ambito di tessuti. Nei vegetali unicellulari in cui non vi è differenziazione di organi, il corpo dell'organismo è detto tallo. Le piante con cormo sono dette cormofite. Un elemento caratterizzante le cormofite è la presenza di tessuti differenziati per il trasporto di liquidi su lunghe distanze, in particolare la linfa grezza dalle radici alle foglie
• Possiedono capacità metaboliche elevate
• Sono in grado di traslocare i metaboliti prodotti a tutte le parti dell’organismo
• Sono in grado di rispondere al variare delle condizioni ambientali e di attuare strategie di adattamento
• Tutto ciò è reso possibile da un sofisticato sistema di regolazione delle attività della pianta
• Il sistema di regolazione è costituito da ORMONI, prodotte dalla pianta stessa, che vengono traslocati attraverso gli elementi conduttori.
Tutte le funzioni del vegetale sono strettamente coordinate: le piante rispondono alle variazioni dei fattori ambientali fra cui luce, gravità, temperatura … modificando la produzione di ormoni (regolatori di crescita) che di conseguenza inducono cambiamenti alle caratteristiche e ai ritmi di crescita vegetale. Principali caratteristiche degli ormoni vegetali
sono molecole organiche a basso peso molecolare sono attive a concentrazioni molto basse un ormone vegetale può essere prodotto in organi differenti nella stessa pianta possono agire anche nello stesso sito di produzione anche se un ormone influisce sull’attività di un altro, non esiste un centro unico di controllo dell’intero sistema ormonale gli ormoni vegetali sono meno numerosi di quelli animali, spesso un ormone agisce su più organi e su più funzioni la stessa quantità di un ormone può determinare risposte diverse in organi differenti uno stesso organo può rispondere in modo differente a seconda della concentrazione del medesimo ormone
GLI ORMONI VEGETALI
I fitormoni sono composti determinanti nell’orientare e regolare la differenziazione e lo sviluppo degli organismi vegetali: hanno funzione di stimolare e di inibire.
Sono in grado di esercitare stimoli specifici sulle diverse funzioni fisiologiche delle piante. Il controllo ormonale si estende sulle principali attività vegetali: germinazione, differenziazione cellulare, accrescimento, lignificazione, fioritura, fruttificazione, dormienza ...
Sono prodotti in determinati distretti della pianta, dove possono agire direttamente o vengono traslocati in altri organi dove condizionano le attività metaboliche delle cellule e quindi lo sviluppo dell'intero organismo
Non tutte le cellule vegetali in coltura sono in grado di produrre ormoni; la produzione è riservata a quelle cellule che sono in grado di attivare la trascrizione e la traduzione dell'informazione genetica che codifica per quei composti.
Agli ormoni viene affidata la capacità della pianta a rispondere agli stimoli ambientali, attraverso una modificazione della crescita della pianta detta tropismo.
Differenza
tra
ormoni
anima Differenza tra ormoni animali e vegetali
Tutti gli ormoni hanno effetto a basse concentrazione. Aumentando la concentrazione aumenta anche l’effetto fino ad arrivare ad una concentrazione ottimale alla quale c’è l’effetto massimo. Tuttavia negli animali l’azione ormonale può essere graduata con piccole variazioni di concentrazione, nelle piante invece sono necessarie variazioni dell’ordine di 1000-10000 volte.
Non vi sono in una pianta organi specializzati a produrre ormoni: i centri di produzione sono multipli e difficili da localizzare
Gli ormoni vegetali possono anche agire sullo stesso sito di produzione
Ciascun ormone vegetale ha una vasta gamma di attività ed agisce su più organi con diverse funzioni; di conseguenza ogni organo vegetale è regolato da più ormoni
L’azione di un ormone vegetale ha effetti differenti nei vari organi e dipende dalla interazione con i diversi recettori che innescano catene di trasduzione differenti.
Gli ormoni animali chimicamente sono polipeptidi o steroidi, quelli vegetali sono molecole più sintetizzate da diverse vie metaboliche e hanno perciò varie strutture.
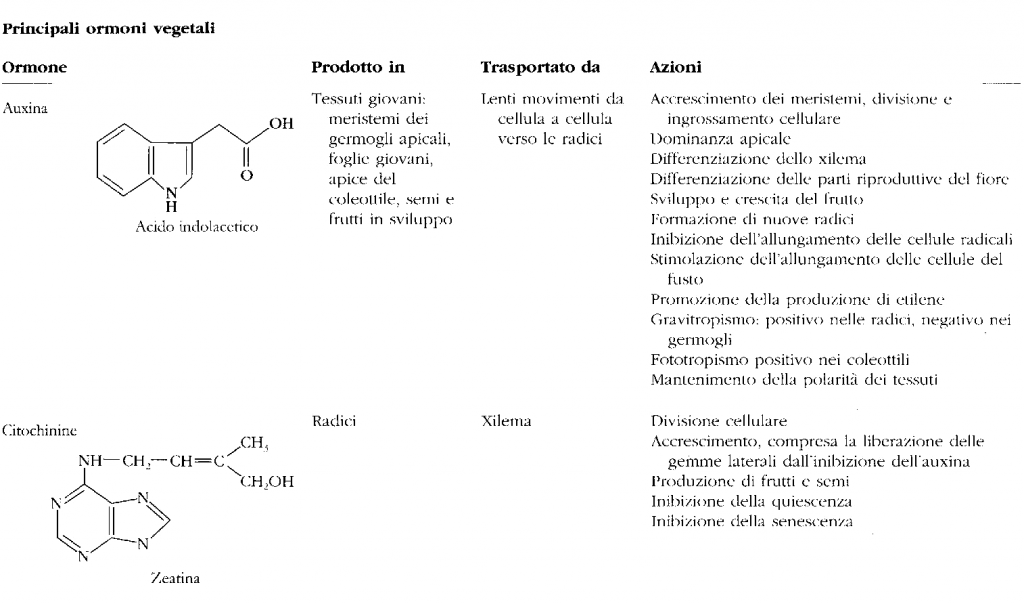
Le piante hanno un numero inferiore di ormoni, i principali sono infatti soltanto cinque: auxine, gibberelline, citochinine, acido abscissico ed etilene.
Negli animali l’azione degli ormoni è regolata da un sistema rigido: la maggior parte delle ghiandole endocrine è regolata dall’ipofisi, che è controllata dal sistema nervoso. Nelle piante il coordinamento è invece meno gerarchico e non esiste un unico centro che comanda e dirige la produzione e la secrezione di tutti gli ormoni. Auxine sono dei composti che hanno in comune la presenza di un gruppo chimico che l'indolo. Le auxine agiscono sui processi di accrescimento e allungamento della pianta. Le principali azioni delle auxine sono di stimolo nei processi: • divisione cellulare • accrescimento cellulare • produzione di nuove radici • produzione di vasi • partenocarpia (sviluppo del frutto senza fecondazione).
Gibberelline Il più importante è l'acido gibberellico. • Agiscono in sinergia con le auxine nella stimolazione della moltiplicazione ed estensione cellulare e nell'induzione della partenocarpa. • Stimola l'accrescimento dei frutti e lo sviluppo delle gemme.
Citochinine Questi ormoni gruppo provocano la divisione cellulare. Agiscono in sinergia con le auxine negli organi in rapido accrescimento.
Acido abscissico Ormone ad azione inibente, sintetizzato soprattutto nelle lami |
Acido abscissico
Ormone ad azione inibente, sintetizzato soprattutto nelle lamine fogliari,
la cui azione più significativa riguarda la dormienza dei semi e delle gemme. Interviene anche nel processo di caduta delle foglie e dei frutti.
Etilene
E' un ormone inibitore e favorisce la crescita e la maturazione dei frutti. Agisce anche come attivatore della germinazione dei semi e nella schiusura
AUXINE


Sono state le prime sostanze regolatrici scoperte nel 1926
• La principale auxina naturale è l’acido indolacetico (IAA), con struttura chimica simile all’aminoacido triptofano
• L’auxina viene prodotta principalmente dai meristemi apicali dei germogli, ed è trasportata nel resto della pianta con un movimento unidirezionale, cioè dal germoglio fino alle radici (può essere prodotta anche negli ovari e nei semi).
• La sostanza è localizzata nei meristemi, negli embrioni, in foglie e frutti giovani
• I tessuti trasportatori sono prevalentemente i parenchimi corticale, midollare e il parenchima floematico
• L’auxina esercita una varietà di effetti sull’accrescimento e sulla morfogenesi vegetale
Stimola la crescita delle cellule per distensione
Auxina aumenta la plasticità della parete favorendo la degradazione dei polimeri che la formano.
Il meccanismo d’azione sembra influenzato dalla auxina che induce l’espulsione dalla cellula di ioni H+ che porta ad un abbassamento del il pH della parete (fino 4.8).
Un ambiente così acido destabilizza e indebolisce i
legami di idrogeno tra le fibrille di cellulosa e le emicellulose.
In tal modo sarebbero attivati enzimi idrolitici che degradano i polisaccaridi, rendendo la parete meno rigida e facilmente “rimodellabile”.
Interviene nei fenomeni di Tropismo
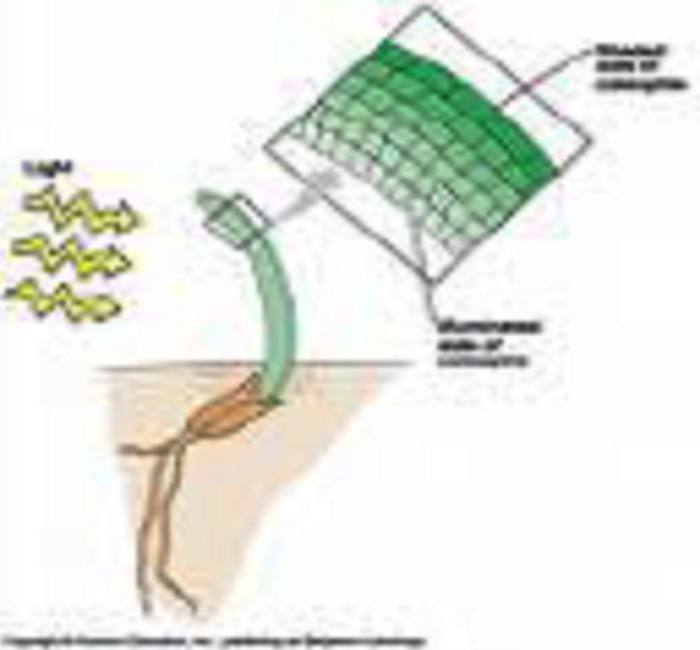
Le auxine hanno un ruolo importante nei fototropismi e geotropismi come conseguenza del processo di distensione che induce l’allungamento delle cellule.
Nel caso del fototropismo a causa della luce l’auxina migra dalla parte illuminata a quella oscura dell’apice, dove promuoverà l’allungamento più rapido delle cellule del lato oscuro rispetto a quello del lato illuminato, per cui la pianta si piegherà verso la luce.
Crescita delle radici:
Le auxine a concentrazioni diverse possono indurre effetti contrastanti:
- in piccole quantità 10-8 – 10-6 M l’auxina stimola la crescita delle radici, anche di quelle laterali e di quelle avventizie.
- ad alte concentrazioni dell’auxina inibiscono la crescita delle radici
Questo effetto dell’auxina sull’iniziazione di radici è utilizzato in agricoltura propagazione di piante per talea.
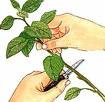

Dominanza apicale:
le auxine prodotte dalla gemma apicale inibiscono o rallentano lo sviluppo delle gemme laterali o secondarie
L’auxina è responsabile della dominanza apicale, poiché, prodotta nell’apice del germoglio, scende lungo il fusto e blocca la crescita delle gemme ascellari.
Infatti se si recide l’apice del germoglio, le gemme
ascellari crescono rapidamente.
Il fenomeno della dominanza apicale è legato:
- sia alla quantità di auxina che giunge dalla gemma apicale,
- sia ad un’azione indiretta dell’auxina che stimola, in prossimità delle gemme laterali, lo sviluppo di un ormone antagonista che ne inibisce l’accrescimento
La presenza di gemme laterali quiescenti è fondamentale per la sopravvivenza in caso di danneggiamento della gemma apicale
Sulla conoscenza di questi fenomeni si basano le pratiche di potatura
Inibizione della caduta delle foglie
Giovani foglie producono auxine e il loro invecchiamento e distacco è correlato ad una diminuzione del contenuto di auxine nella pianta (ed ad una stimolazione dell’Etilene).
L’IAA è in grado di ritardare i primi stadi di abscissione fogliare: le concentrazioni di auxina sono alte nelle giovani foglie, diminuiscono progressivamente nelle foglie mature e sono relativamente basse nelle foglie senescenti.
Ripresa
Ha un ruolo crescita sta a produrre x
• Con la rip meristemi p
le divisioni cellulari del cambio cribro-vascolare.
• Le nuove foglie prodotte sono collegate al nuovo tessuto conduttore.
• Anche l’attività del fellogeno viene stimolata
• L’auxina favorisce anche la produzione di tessuti cicatriziali in caso di ferite
Regolazione dello sviluppo dei frutti
La auxina è anche responsabile della crescita dei frutti. In particolare se viene posta a contatto con un ovario di un fiore lo fa trasformare in un frutto, che però non
contiene semi in quanto non è avvenuta una fecondazione.
La produzione di frutti ottenuta senza fecondazione è definita partenocarpia ed è sfruttata in agricoltura.

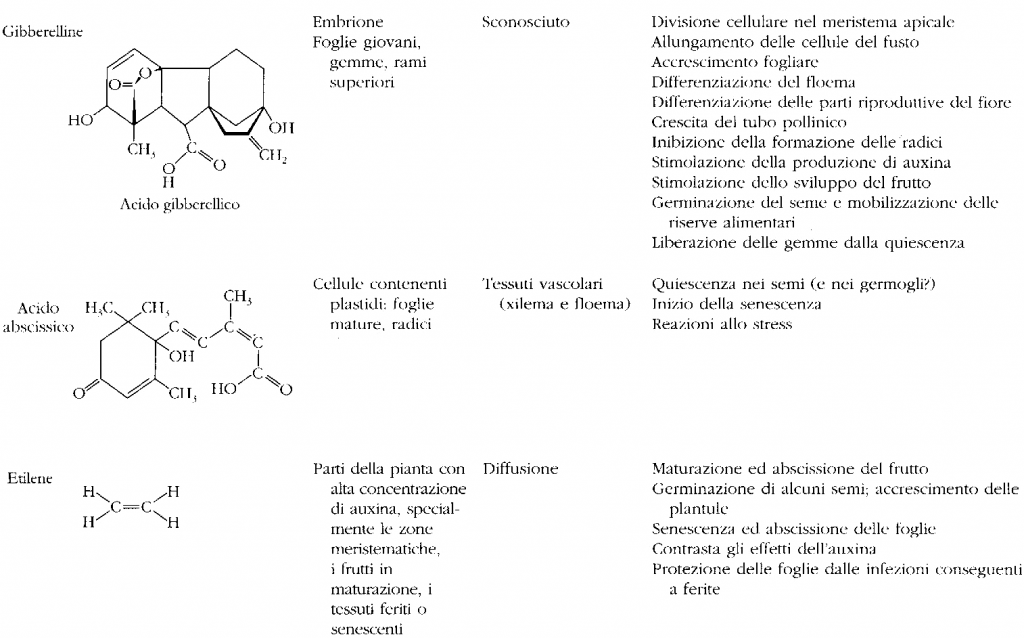
GIBBERELLINE
• Attualmente sono state identificate circa 70 tipi di gibberelline.
• Hanno una complessa struttura chimica terpenica ciclica (diterpeni);
mostrano lievi differenze strutturali e possono essere convertite una nell’altra
• La struttura complessa rende difficile ottenere analog
• La gibberellina più diffusa è l’acido gibberellico (GA)
• Sono sintetizzate nei meristemi del fusto e della radice, nelle foglie giovani e nei frutti immaturi
• Sono distribuite in tutti gli organi della pianta;
i frutti immaturi ne contengono quantità elevate
• Il loro trasporto avviene attraverso il floema dai semi alla plantula; attraverso il legno nella radice; dalle foglie agli altri organi attraverso il parenchima, in modo apolare.
• Il loro meccanismo di azione, nei semi, sembra che si realizzi mediante il controllo della sintesi dell’RNA
E’ stata isolata per la prima volta nel 1926 in Giappone da un fungo parassita del riso (Gibberella fujikuroi).
Il fungo producendo la gibberellina determinava un allungamento abnorme del fusto e delle foglie;
le piante crescevano deboli e stentate.
I luoghi di produzione di gibberelline sono:
- le gemme,
- le foglie giovani
- semi
Foglie di Arabidopsis con e senza GA
Stimolazione del processo di divisione e distensione cellulare, che provoca un allungamento del fusto. L’accrescimento è stimolato dalla presenza di entrambi gli ormoni: gibberelline e auxine che agiscono in maniera sinergica.
Le gibberelline non inibiscono l’accrescimento delle radici neppure ad elevate concentrazioni

Stimolazione della fioritura: le gibberelline inducono il passaggio dalla fase vegetativa a quella riproduttiva della pianta. Se si applica una soluzione molto diluita a base di gibberelline sulla gemma apicale, dapprima la pianta cresce e si allunga, poi incomincia a fiorire.
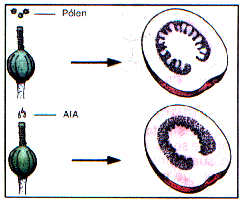
Stimolazione della germinazione del polline e successiva formazione del tubetto pollinico.
Promozione nello sviluppo dei frutti ed in particolar modo di di frutti partenocarpici (mele, cocomeri, cetriolo)
Riattivazione del cambio cribro- vascolare e la produzione di
floema secondario
Mantengono lo stadio giovanile ritardando la senescenza di foglie e frutti
Interrompono la quiescenza dei semi: dopo l’imbibizione l’embrione libera gibberelline che stimolano le cellule sotto il tegumento a sintetizzare gli enzimi idrolitici per la degradazione delle sostanze di riserva.
Le gibberelline vengono prodotte anche nell’embrione presente nel seme immaturo.
Questi ormoni diffonderebbero nello strato aleuronico, ricco di amidi e proteine, e qui promuoverebbero la sintesi di enzimi in grado di idrolizzare lipidi, proteine e zuccheri per permettere all’embrione di ricavare energia e sostanze utili per il proprio metabolismo.
CITOCHININE
Le citochinine sono ormoni che stimolano la divisione cellulare e si trovano nei meristemi, nei semi in germinazione, nei frutti e nelle radici.
Il primo composto ad azione citochininica fu la zeatina, isolato per la prima volta nel 1964 dal mais (Zea mais). Ad oggi sono state identificate più di 40 specie di citochinine.
• Fra quelle naturali la più attiva è la zeatina, mentre tra quelle sintetiche è molto attiva la cinetina
• Hanno struttura chimica simile alla base azotata
adenina, da cui derivano
• Sono sintetizzate nell’apice radicale
• Sono presenti in tutti i tessuti giovani, in attiva divisione. Particolarmente abbondanti nei semi, negli apici radicali, ma anche in foglie e frutti giovani
• Vengono traslocate attraverso il legno dalle radici alle altre parti del vegetale
Stimolano la divisione cellulare Stimolano l’accrescimento dei cotiledoni, aumentando la plasticità delle pareti cellulari. In fusti e radici la somministrazione di citochinine tende a far allargare le cellule (forse per cambiamento di orientamento delle microfibrille di cellulosa di neoformazione ).
Promuovono la maturazione dei cloroplasti Promuovono la sintesi di proteine fotosintetiche: foglie eziolate trattate con citochinine prima di essere illuminate formano cloroplasti con grana più estesi.
Le clorofille e gli enzimi fotosintetici vengono anche sintetizzati a velocità maggiore dopo l’illuminazione.
Stimolano la mobilitazione delle sostanze nutrienti L’ormone stimola la mobilitazione delle sostanze nutritive, quindi in successione ciò porta alla attivazione metabolica dell’area trattata.
In particolare durante la germinazione viene stimolato il passaggio delle sostanze nutritive dall’organo di riserva (seme) alla foglia per innescare la attività fotosintetica.
Ritardano la senescenza delle foglie
L’effetto di ritardare la senescenza delle foglie può essere evidenziato confrontando foglie trattate e non trattate con citochinine.
In particolare le citochinine giocano un ruolo importante sull’accrescimento generale
delle piante in combinazione sinergica con l’auxina
Regolano il ciclo cellulare vegetale
Le citochinine innescano la proliferazione cellulare in tessuti che contengono, o a cui si è aggiunta, una concentrazione ottimale di auxine.
Entrambi gli ormoni partecipano alla regolazione del ciclo cellulare:
• auxina regola gli eventi che portano alla replicazione del DNA,
• citochinine regolano gli eventi che portano alla mitosi.
Il rapporto Auxina/Citochinina regola la morfogenesi nelle colture di tessuti
Esperimenti hanno dimostrato che i due ormoni regolano la formazione degli organi:
- alte concentrazioni di auxina: stimolavano la formazione di radici,
e: inducono la formazione di gemme e germogli. uti crescevano come calli indifferenziati.
tessuto calloso
Nelle colture in vitro, l’aggiunta nel mezzo di coltura della sola auxina determina un aumento delle dimensioni cellulari, ma se insieme all’auxina si aggiungono delle citochinine si assiste ad un rapido incremento della divisione cellulare, con formazione di numerose ma piccole cellule indifferenziate.
Quindi quando i due ormoni sono a concentrazioni uguali, le cellule restano indifferenziate e formano una massa di tessuto detto CALLO.

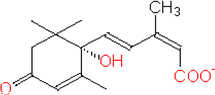
ACIDO ABSCISSICO
- Isolato nel 1960; non sono ancora chiari i siti di produzione, ma si accumula nelle foglie e nei frutti poco prima del distacco, nelle gemme e nei semi quiescenti.
- E’ considerato un ormone inibitorio, in quanto ABA è il diretto antagonista degli ormoni detti della crescita (auxine, gibberelline, citochinine).
- È presente nei plastidi (cloroplasti di foglie mature), nei semi e nelle radici. Risulta quindi essere un ormone ubiquitario poiché è sintetizzato in tutte le cellule che contengono cloroplasti o amiloplasti.
- Biogenesi da acido mevalonico (chimicamente appartiene ai sesquiterpeni, derivati dall’isoprene); prodotto di degradazione di carotenoidi e xantofille.

Regola l’abscissione e la senescenza
L’ABA con etilene è responsabile del fenomeno dell’abscissione, che si manifesta con la caduta delle foglie, o il distacco in un frutto. Alla base del picciolo ABA induce la proliferazione di cellule piccole, piatte con pareti molto sottili e prive di sostegno meccanico: strato di abscissione.
Le lamelle mediane gelificano lasciando la foglia attaccata al ramo solo per mezzo dei fasci, che finiscono per rompersi provocando la caduta della foglia.
Al di sotto dello strato di abscissione si forma uno strato protettivo con cellule suberificate, in grado di isolare la foglia dal fusto prima della caduta.
Inibisce l’accrescimento della pianta
L’accrescimento delle piante indotto dall’auxina viene inibito dall’ABA, che per questo motivo viene definito ormone inibitor di crescita. L’ABA blocca l’estrusione di H+ da parte dell’auxin prevenendo quindi l’acidificazione della parete cellulare e la distensione della cellula.
Risposta agli stress e chiusura degli stomi
L’ABA è stato definito ormone da stress essendo implicato nella risposta a stimoli esterni come freddo, salinità e umidità. In risposta a stress idrico, ABA induce la chiusura degli stomi riducendo la perdita di H2O dovuta a traspirazione.
Regola l’assorbimento di acqua
Nei tessuti radicali ABA stimola l’assorbimento idrico e quello ionico, aumentando così, in associazione con la chiusura degli stomi,
il turgore della pianta.
Stimola l’accrescimento radicale
l’ABA induce l’accrescimento radicale e stimola la fuoriuscita di radici laterali, sopprimendo allo stesso temp
Questi effetti antagonistici dell la riduzione dell’area fogliare all’assorbimento dell’acqua, tu con le condizioni di siccità.
Regola la dormienza delle gemme
Nei climi freddi la dormienza è un’importante caratteristica adattativa. Quando durante l’inverno un albero affronta temperature molto fredde protegge i suoi meristemi con le perule e interrompe temporaneamente l’accrescimento della gemma..
Le interazioni fra l’ABA e altri ormoni sono responsabili di un processo in cui la dormienza della gemma e l’accrescimento sono regolati dal bilancio fra gli
inibitori di accrescimento, come l’ABA, e le sostanze
che inducono la crescita, ad esempio le citochinine e
gibberelline.
Regola la dormienza dei semi
I semi dormienti contengono di solito concentrazioni superiori di ABA rispetto a quelli non dormienti. Come nel caso della dormienza delle gemme, è il bilancio ormonale (più che le fluttuazioni della concentrazione di un singolo ormone) che risulta controllare la transizione dalla dormienza alla germinazione.
L’ABA inibisce la sintesi di enzimi idrolitici che sono fondamentali per la degradazione di sostanze di riserva del seme che quindi si mantiene nel suo status di dormienza, inibendo in tal modo la germinazione
• L’etilene è un ormone biologicamente attivo a concentrazioni bassissime, ed essendo un gas la sua azione si trasmette anche a distanza per diffusione.
• L’etilene si forma a partire dall’a.a. metionina, i fattori di controllo che intervengono nella sintesi di etilene sono principalmente le condizioni ambientali (O2, T°, luce). L’attività dell’etilene aumenta all’aumentare della concentrazione di ossigeno e diminuisce man mano che l’ambiente si satura di anidride carbonica.
• L’etilene è responsabile della caduta delle foglie e della maturazione dei frutti, del cambiamento del loro colore e della loro consistenza e composizione chimica.
• Le più alte produzioni di etilene avvengono in tessuti senescenti o in frutti in via di maturazione. Viene sintetizzato in quantità e in distretti diversi a seconda dell'età della pianta
• Rispetto alla risposta delle piante alla gravità, l’etilene agisce in maniera contraria all’auxina: semi esposti ad etilene, manifestano un accrescimento orizzontale delle radici durante la germinazione.
ETILENE
Regola l’abscissione e i fenomeni di invecchiamento L’etilene, in azione sinergica con acido abscissico, è responsabile dell’ingiallimento, appassimento e caduta delle foglie,
fiori, frutti e altri organi vegetali.
Causa l'indebolimento delle pareti cellulari dello strato di abscissione, promuovendo l’azione di enzimi (cellulasi e poligalatturasi) che degradano la parete cellulare.
Maturazione dei frutti
L’etilene promuove la maturazione di molti frutti, dopo che essi si sono sviluppati in seguito a stimoli di auxina, gibberelline e citochinine.
La maturazione di un frutto comporta:
• aumento della velocità di respirazione,
• accumulo di zuccheri solubili,
• idrolisi parziale delle pareti cellulari del parenchima che forma la polpa,
• sintesi di molecole aromatiche che danno profumo al frutto.

Inibizione della divisione cellulare e dell’allungamento
L’etilene agisce in maniera contraria rispetto all’auxina:
• inibisce la divisione cellulare sia nelle radici sia nel fusto;
• blocca il processo mitotico attraverso l'inibizione della sintesi di DNA
• inibisce l'allungamento cellulare per distensione
• altera la direzione dell’accrescimento cellulare: le cellule si es tutte le direzioni, anziché allungarsi in senso prevalentemente l
Abolizione del geotropismo positivo delle radici
• Rispetto alla risposta delle piante alla gravità, l’etilene agisce i l’auxina: semi esposti ad etilene, manifestano un accresciment durante la germinazione.
• E’ responsabile della curvatura ad uncino dell'apice vegetativ nello sviluppo sotterraneo del germinello del seme.
• L'etilene cambia i modelli di accrescimento delle plantule riduc la velocità di allungamento e aumentando l'espansione laterale, portando così al rigonfiamento della zona posta sotto la parte
a forma di gancio.
Inibizione dormienza semi e gemme
l’etilene quando è applicato a semi di cereali interrompe la dor e dà inizio alla germinazione, regolandone anche la velocità.